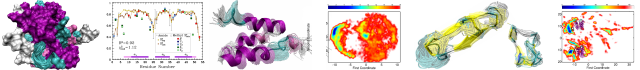

Journal Articles
Discrimination of Kinetic Models by a Combination of Microirradiation and Fluorescence Photobleaching
Secondary Structure Analysis of a Functional Construct of Caveolin-1 Reveals a Long C-Terminal Helix
The Effect of Membrane Lipid Composition on the Formation of Lipid Ultrananodomains
Can Specific Protein-Lipid Interactions Stabilize an Active State of the Beta 2 Adrenergic Receptor?
Mapping the Processivity Determinants of the Kinesin-3 Motor Domain
Theory of Triplet Excitation Transfer in the Donor-Oxygen-Acceptor System: Application to Cytochrome f
Active Biochemical Regulation of Cell Volume and a Simple Model of Cell Tension Response
Critical Timing without a Timer for Embryonic Development
Local Crystalline Structure in an Amorphous Protein Dense Phase
Dipole-Potential-Mediated Effects on Ion Pump Kinetics
A Thermodynamic Model for Genome Packaging in Hepatitis B Virus
MDTraj: A Modern Open Library for the Analysis of Molecular Dynamics Trajectories
Nucleosome Core Particle Disassembly and Assembly Kinetics Studied Using Single-Molecule Fluorescence
Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract
Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract
Nature 526, 7575 (2015). doi:10.1038/nature15524
Authors: Sushma Kommineni, Daniel J. Bretl, Vy Lam, Rajrupa Chakraborty, Michael Hayward, Pippa Simpson, Yumei Cao, Pavlos Bousounis, Christopher J. Kristich & Nita H. Salzman
Enterococcus faecalis is both a common commensal of the human gastrointestinal tract and a leading cause of hospital-acquired infections. Systemic infections with multidrug-resistant enterococci occur subsequent to gastrointestinal colonization. Preventing colonization by multidrug-resistant E. faecalis could therefore be a valuable approach towards limiting infection. However, little is known about the mechanisms E. faecalis uses to colonize and compete for stable gastrointestinal niches. Pheromone-responsive conjugative plasmids encoding bacteriocins are common among enterococcal strains and could modulate niche competition among enterococci or between enterococci and the intestinal microbiota. We developed a model of colonization of the mouse gut with E. faecalis, without disrupting the microbiota, to evaluate the role of the conjugative plasmid pPD1 expressing bacteriocin 21 (ref. 4) in enterococcal colonization. Here we show that E. faecalis harbouring pPD1 replaces indigenous enterococci and outcompetes E. faecalis lacking pPD1. Furthermore, in the intestine, pPD1 is transferred to other E. faecalis strains by conjugation, enhancing their survival. Colonization with an E. faecalis strain carrying a conjugation-defective pPD1 mutant subsequently resulted in clearance of vancomycin-resistant enterococci, without plasmid transfer. Therefore, bacteriocin expression by commensal bacteria can influence niche competition in the gastrointestinal tract, and bacteriocins, delivered by commensals that occupy a precise intestinal bacterial niche, may be an effective therapeutic approach to specifically eliminate intestinal colonization by multidrug-resistant bacteria, without profound disruption of the indigenous microbiota.
An Optimal Algorithm for the Weighted Backup 2-Center Problem on a Tree
In this paper, we are concerned with the weighted backup 2-center problem on a tree. The backup 2-center problem is a kind of center facility location problem, in which one is asked to deploy two facilities, with a given probability to fail, in a network. Given that the two facilities do not fail simultaneously, the goal is to find two locations, possibly on edges, that minimize the expected value of the maximum distance over all vertices to their closest functioning facility. In the weighted setting, each vertex in the network is associated with a nonnegative weight, and the distance from vertex u to v is weighted by the weight of u. With the strategy of prune-and-search, we propose a linear time algorithm, which is asymptotically optimal, to solve the weighted backup 2-center problem on a tree.
Improved Subquadratic 3SUM
In the 3SUM problem we are given three lists \(\mathcal {A}\) , \(\mathcal {B}\) , \(\mathcal {C}\) , of n real numbers, and are asked to find \((a,b,c)\in \mathcal {A}\times \mathcal {B}\times \mathcal {C}\) such that \(a+b=c\) . The longstanding 3SUM conjecture—that 3SUM could not be solved in subquadratic time—was recently refuted by Grønlund and Pettie. They presented \(\hbox {O}\left( n^2(\log \log n)^{\alpha }/(\log n)^{\beta }\right) \) algorithms for 3SUM and for the related problems Convolution3SUM and ZeroTriangle, where \(\alpha \) and \(\beta \) are constants that depend on the problem and whether the algorithm is deterministic or randomized (and for ZeroTriangle the main factor is \(n^3\) rather than \(n^2\) ). We simplify Grønlund and Pettie’s algorithms and obtain better bounds, namely, \(\alpha =\beta =1\) , deterministically. For 3SUM our bound is better than both the deterministic and the randomized bounds of Grønlund and Pettie. For the other problems our bounds are better than their deterministic bounds, and match their randomized bounds.
Synergistic Catalysis of Ionic Brønsted Acid and Photosensitizer for a Redox Neutral Asymmetric α-Coupling of N-Arylaminomethanes with Aldimines
Large-Scale Conformational Dynamics Control H5N1 Influenza Polymerase PB2 Binding to Importin α
Crystal structures of halohydrin hydrogen-halide-lyases from Corynebacterium sp. N-1074
Halohydrin hydrogen-halide-lyase (H-Lyase) is a bacterial enzyme that is involved in the degradation of halohydrins. This enzyme catalyzes the intramolecular nucleophilic displacement of a halogen by a vicinal hydroxyl group in halohydrins to produce the corresponding epoxides. The epoxide products are subsequently hydrolyzed by an epoxide hydrolase, yielding the corresponding 1, 2-diol. Until now, six different H-Lyases have been studied. These H-Lyases are grouped into three subtypes (A, B, and C) based on amino acid sequence similarities and exhibit different enantioselectivity. Corynebacterium sp. strain N-1074 has two different isozymes of H-Lyase, HheA (A-type) and HheB (B-type). We have determined their crystal structures to elucidate the differences in enantioselectivity among them. All three groups share a similar structure, including catalytic sites. The lack of enantioselectivity of HheA seems to be due to the relatively wide size of the substrate tunnel compared to that of other H-Lyases. Among the B-type H-Lyases, HheB shows relatively high enantioselectivity compared to that of HheBGP1. This difference seems to be due to amino acid replacements at the active site tunnel. The binding mode of 1, 3-dicyano-2-propanol at the catalytic site in the crystal structure of the HheB-DiCN complex suggests that the product should be (R)-epichlorohydrin, which agrees with the enantioselectivity of HheB. Comparison with the structure of HheC provides a clue for the difference in their enantioselectivity. Proteins 2015; 83:2230–2239. © 2015 Wiley Periodicals, Inc.
pKa predictions for proteins, RNAs, and DNAs with the Gaussian dielectric function using DelPhi pKa
We developed a Poisson-Boltzmann based approach to calculate the values of protein ionizable residues (Glu, Asp, His, Lys and Arg), nucleotides of RNA and single stranded DNA. Two novel features were utilized: the dielectric properties of the macromolecules and water phase were modeled via the smooth Gaussian-based dielectric function in DelPhi and the corresponding electrostatic energies were calculated without defining the molecular surface. We tested the algorithm by calculating values for more than 300 residues from 32 proteins from the PPD dataset and achieved an overall RMSD of 0.77. Particularly, the RMSD of 0.55 was achieved for surface residues, while the RMSD of 1.1 for buried residues. The approach was also found capable of capturing the large shifts of various single point mutations in staphylococcal nuclease (SNase) from -cooperative dataset, resulting in an overall RMSD of 1.6 for this set of pKa's. Investigations showed that predictions for most of buried mutant residues of SNase could be improved by using higher dielectric constant values. Furthermore, an option to generate different hydrogen positions also improves predictions for buried carboxyl residues. Finally, the calculations on two RNAs demonstrated the capability of this approach for other types of biomolecules. Proteins 2015; 83:2186–2197. © 2015 Wiley Periodicals, Inc.