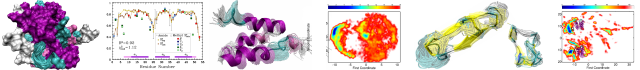

Journal Articles
Electric-Field-Driven Dual Vacancies Evolution in Ultrathin Nanosheets Realizing Reversible Semiconductor to Half-Metal Transition
Protein structure determination by conformational space annealing using NMR geometric restraints
We have carried out numerical experiments to investigate the applicability of the global optimization method of conformational space annealing (CSA) to the enhanced NMR protein structure determination over existing PDB structures. The NMR protein structure determination is driven by the optimization of collective multiple restraints arising from experimental data and the basic stereochemical properties of a protein-like molecule. By rigorous and straightforward application of CSA to the identical NMR experimental data used to generate existing PDB structures, we redetermined 56 recent PDB protein structures starting from fully randomized structures. The quality of CSA-generated structures and existing PDB structures were assessed by multiobjective functions in terms of their consistencies with experimental data and the requirements of protein-like stereochemistry. In 54 out of 56 cases, CSA-generated structures were better than existing PDB structures in the Pareto-dominant manner, while in the remaining two cases, it was a tie with mixed results. As a whole, all structural features tested improved in a statistically meaningful manner. The most improved feature was the Ramachandran favored portion of backbone torsion angles with about 8.6% improvement from 88.9% to 97.5% (P-value <10−17). We show that by straightforward application of CSA to the efficient global optimization of an energy function, NMR structures will be of better quality than existing PDB structures. Proteins 2015; 83:2251–2262. © 2015 Wiley Periodicals, Inc.
New encouraging developments in contact prediction: Assessment of the CASP11 results
This article provides a report on the state-of-the-art in the prediction of intra-molecular residue-residue contacts in proteins based on the assessment of the predictions submitted to the CASP11 experiment. The assessment emphasis is placed on the accuracy in predicting long-range contacts. Twenty-nine groups participated in contact prediction in CASP11. At least eight of them used the recently developed evolutionary coupling techniques, with the top group (CONSIP2) reaching precision of 27% on target proteins that could not be modeled by homology. This result indicates a breakthrough in the development of methods based on the correlated mutation approach. Successful prediction of contacts was shown to be practically helpful in modeling three-dimensional structures; in particular target T0806 was modeled exceedingly well with accuracy not yet seen for ab initio targets of this size (>250 residues). Proteins 2015. © 2015 Wiley Periodicals, Inc.
Template-free modeling by LEE and LEER in CASP11
For the template-free modeling of human targets of CASP11, we utilized two of our modeling protocols, LEE and LEER. The LEE protocol took CASP11-released server models as the input and used some of them as templates for 3D (three-dimensional) modeling. The template selection procedure was based on the clustering of the server models aided by a community detection method of a server-model network. Restraining energy terms generated from the selected templates together with physical and statistical energy terms were used to build 3D models. Side-chains of the 3D models were rebuilt using target-specific consensus side-chain library along with the SCWRL4 rotamer library, which completed the LEE protocol. The first success factor of the LEE protocol was due to efficient server model screening. The average backbone accuracy of selected server models was similar to that of top 30% server models. The second factor was that a proper energy function along with our optimization method guided us, so that we successfully generated better quality models than the input template models. In 10 out of 24 cases, better backbone structures than the best of input template structures were generated. LEE models were further refined by performing restrained molecular dynamics simulations to generate LEER models. CASP11 results indicate that LEE models were better than the average template models in terms of both backbone structures and side-chain orientations. LEER models were of improved physical realism and stereo-chemistry compared to LEE models, and they were comparable to LEE models in the backbone accuracy. Proteins 2015. © 2015 Wiley Periodicals, Inc.
Partial unfolding and refolding for structure refinement: A unified approach of geometric simulations and molecular dynamics
The most successful protein structure prediction methods to date have been template-based modeling (TBM) or homology modeling, which predicts protein structure based on experimental structures. These high accuracy predictions sometimes retain structural errors due to incorrect templates or a lack of accurate templates in the case of low sequence similarity, making these structures inadequate in drug-design studies or molecular dynamics simulations. We have developed a new physics based approach to the protein refinement problem by mimicking the mechanism of chaperons that rehabilitate misfolded proteins. The template structure is unfolded by selectively (targeted) pulling on different portions of the protein using the geometric based technique FRODA, and then refolded using hierarchically restrained replica exchange molecular dynamics simulations (hr-REMD). FRODA unfolding is used to create a diverse set of topologies for surveying near native-like structures from a template and to provide a set of persistent contacts to be employed during re-folding. We have tested our approach on 13 previous CASP targets and observed that this method of folding an ensemble of partially unfolded structures, through the hierarchical addition of contact restraints (that is, first local and then nonlocal interactions), leads to a refolding of the structure along with refinement in most cases (12/13). Although this approach yields refined models through advancement in sampling, the task of blind selection of the best refined models still needs to be solved. Overall, the method can be useful for improved sampling for low resolution models where certain of the portions of the structure are incorrectly modeled. Proteins 2015; 83:2279–2292. © 2015 Wiley Periodicals, Inc.
Amino acid positions subject to multiple coevolutionary constraints can be robustly identified by their eigenvector network centrality scores
As proteins evolve, amino acid positions key to protein structure or function are subject to mutational constraints. These positions can be detected by analyzing sequence families for amino acid conservation or for coevolution between pairs of positions. Coevolutionary scores are usually rank-ordered and thresholded to reveal the top pairwise scores, but they also can be treated as weighted networks. Here, we used network analyses to bypass a major complication of coevolution studies: For a given sequence alignment, alternative algorithms usually identify different, top pairwise scores. We reconciled results from five commonly-used, mathematically divergent algorithms (ELSC, McBASC, OMES, SCA, and ZNMI), using the LacI/GalR and 1,6-bisphosphate aldolase protein families as models. Calculations used unthresholded coevolution scores from which column-specific properties such as sequence entropy and random noise were subtracted; “central” positions were identified by calculating various network centrality scores. When compared among algorithms, network centrality methods, particularly eigenvector centrality, showed markedly better agreement than comparisons of the top pairwise scores. Positions with large centrality scores occurred at key structural locations and/or were functionally sensitive to mutations. Further, the top central positions often differed from those with top pairwise coevolution scores: instead of a few strong scores, central positions often had multiple, moderate scores. We conclude that eigenvector centrality calculations reveal a robust evolutionary pattern of constraints—detectable by divergent algorithms—that occur at key protein locations. Finally, we discuss the fact that multiple patterns coexist in evolutionary data that, together, give rise to emergent protein functions. Proteins 2015; 83:2293–2306. © 2015 Wiley Periodicals, Inc.
Toward decrypting the allosteric mechanism of the ryanodine receptor based on coarse-grained structural and dynamic modeling
The ryanodine receptors (RyRs) are a family of calcium (Ca) channels that regulate Ca release by undergoing a closed-to-open gating transition in response to action potential or Ca binding. The allosteric mechanism of RyRs gating, which is activated/regulated by ligand/protein binding >200 Å away from the channel gate, remains elusive for the lack of high-resolution structures. Recent solution of the closed-form structures of the RyR1 isoform by cryo-electron microscopy has paved the way for detailed structure-driven studies of RyRs functions. Toward elucidating the allosteric mechanism of RyRs gating, we performed coarse-grained modeling based on the newly solved closed-form structures of RyR1. Our normal mode analysis captured a key mode of collective motions dominating the observed structural variations in RyR1, which features large outward and downward movements of the peripheral domains with the channel remaining closed, and involves hotspot residues that overlap well with key functional sites and disease mutations. In particular, we found a key interaction between a peripheral domain and the Ca-binding EF hand domain, which may allow for direct coupling of Ca binding to the collective motions as captured by the above mode. This key mode was robustly reproduced by the normal mode analysis of the other two closed-form structures of RyR1 solved independently. To elucidate the closed-to-open conformational changes in RyR1 with amino-acid level of details, we flexibly fitted the closed-form structures of RyR1 into a 10-Å cryo-electron microscopy map of the open state. We observed extensive structural changes involving the peripheral domains and the central domains, resulting in the channel pore opening. In sum, our findings have offered unprecedented structural and dynamic insights to the allosteric mechanism of RyR1 via modulation of the key collective motions involved in RyR1 gating. The predicted hotspot residues and open-form conformation of RyR1 will guide future mutational and functional studies. Proteins 2015; 83:2307–2318. © 2015 Wiley Periodicals, Inc.
Strategies to reduce end-product inhibition in family 48 glycoside hydrolases
Family 48 cellobiohydrolases are some of the most abundant glycoside hydrolases in nature. They are able to degrade cellulosic biomass, therefore serve as good enzyme candidates for biofuel production. Family 48 cellulases hydrolyze cellulose chains via a processive mechanism, and produce the end products composed of mainly cellobiose as well as other cellooligomers (dp ≤ 4). The challenge of utilizing cellulases in biofuel production lies in their extremely slow turnover rate. One reason for the low enzyme activity is suggested to be the product binding moderately to enzyme and inhibiting its future performance. In this study, we quantitatively evaluated the product inhibitory effect of four family 48 glycoside hydrolases using molecular dynamics simulations and product expulsion free energy calculations. We also suggested a series of single mutants of the four family 48 glycoside hydrolases with theoretically reduced level of product inhibition. The theoretical calculations provide a guide for future experimental studies on making mutant cellulases with enhanced activity. This article is protected by copyright. All rights reserved.
Molecular dynamics simulations elucidate the mode of protein recognition by Skp1 and the F-box domain in the SCF complex
Polyubiquitination of the target protein by a ubiquitin transferring machinery is key to various cellular processes. E3 ligase Skp1-CUL1-F-box (SCF) is one such complex which plays crucial role in substrate recognition and transfer of the ubiquitin molecule. Previous computational studies have focused on S-phase kinase-associated protein 2 (Skp2), cullin, and RING-finger proteins of this complex, but the roles of the adapter protein Skp1 and F-box domain of Skp2 have not been determined. Using sub-microsecond molecular dynamics simulations of full-length Skp1, unbound Skp2, Skp2-Cks1 (Cks1: Cyclin-dependent kinases regulatory subunit 1), Skp1-Skp2, and Skp1-Skp2-Cks1 complexes, we have elucidated the function of Skp1 and the F-box domain of Skp2. We found that the L16 loop of Skp1, which was deleted in previous X-ray crystallography studies, can offer additional stability to the ternary complex via its interactions with the C-terminal tail of Skp2. Moreover, Skp1 helices H6, H7, and H8 display vivid conformational flexibility when not bound to Skp2, suggesting that these helices can recognize and lock the F-box proteins. Furthermore, we observed that the F-box domain could rotate (3°-123°), and that the binding partner determined the degree of conformational flexibility. Finally, Skp1 and Skp2 were found to execute a domain motion in Skp1-Skp2 and Skp1-Skp2-Cks1 complexes that could decrease the distance between ubiquitination site of the substrate and the ubiquitin molecule by 3nm. Thus, we propose that both the F-box domain of Skp2 and Skp1-Skp2 domain motions displaying preferential conformational control can together facilitate polyubiquitination of a wide variety of substrates. This article is protected by copyright. All rights reserved.
Probing the roles of two tryptophans surrounding the unique zinc coordination site in lipase family I.5
A unique zinc domain found in all of the identified members of the lipase family I.5 is surrounded by two conserved tryptophans (W61 and W212). In this study we investigated the role of these hydrophobic residues in thermostability and thermoactivity of the lipase from Bacillus thermocatenulatus (BTL2) taken as the representative of the family. Circular dichroism spectroscopy revealed that the secondary structure of BTL2 is conserved by the tryptophan mutations (W61A, W212A and W61A/W212A), and that W61 is located in a more rigid and less solvent exposed region than is W212. Thermal denaturation and optimal activity analyses pointed out that zinc induces thermostability and thermoactivity of BTL2, in which both tryptophans W61 and W212 play contributing roles. Molecular explanations describing the roles of these tryptophans were pursued by X-ray crystallography of the open form of the W61A mutant and molecular dynamics simulations which highlighted a critical function for W212 in zinc binding to the coordination site. This study reflects the potential use of hydrophobic amino acids in vicinity of metal coordination sites in lipase biocatalysts design. This article is protected by copyright. All rights reserved.
Stability strengths and weaknesses in protein structures detected by statistical potentials. Application to bovine seminal ribonuclease
We present an in silico method to estimate the contribution of each residue in a protein to its overall stability using three database-derived statistical potentials that are based on inter-residue distances, backbone torsion angles and solvent accessibility, respectively. Residues that contribute very unfavorably to the folding free energy are defined as stability weaknesses, whereas residues that show a highly stabilizing contribution are called stability strengths. Strengths and/or weaknesses on residues that are in spatial contact are clustered into 3-dimensional (3D) stability patches. The identification and analysis of strength- and weakness-containing regions in a protein may reveal its structural or functional characteristics, and/or interesting spots to introduce mutations. To illustrate the power of our method, we apply it to bovine seminal ribonuclease. This enzyme catalyzes the degradation of RNA strands, and has the peculiarity of undergoing 3D domain swapping in physiological conditions. The weaknesses and strengths were compared among the monomeric, dimeric and swapped dimeric forms. We identified weaknesses among the catalytic residues and a mixture of weaknesses and strengths among the substrate-binding residues in the three forms. In the regions involved in 3D swapping, we observed an accumulation of weaknesses in the monomer, which disappear in the dimer and especially in the swapped dimer. Moreover, monomeric homologous proteins were found to exhibit less weaknesses in these regions, whereas mutants known to favor unswapped dimerization appear stabilized in this form. Our method has several perspectives for functional annotation, rational prediction of targeted mutations, and mapping of stability changes upon conformational rearrangements. This article is protected by copyright. All rights reserved.
A composite approach towards a complete model of the myosin rod
Sarcomeric myosins have the remarkable ability to form regular bipolar thick filaments that, together with actin thin filaments, constitute the fundamental contractile unit of skeletal and cardiac muscle. This has been established for over fifty years and yet a molecular model for the thick filament has not been attained. In part this is due to the lack of a detailed molecular model for the coiled-coil that constitutes the myosin rod. The ability to self-assemble resides in the C-terminal of the section of myosin known as light meromyosin (LMM) which exhibits strong salt dependent aggregation that has inhibited structural studies. Here we evaluate the feasibility of generating a complete model for the myosin rod by combining overlapping structures of five sections of coiled-coil covering 164 amino acid residues which constitute 20% of LMM. Each section contains ∼7-9 heptads of myosin. The problem of aggregation was overcome by incorporating the globular folding domains, Gp7 and Xrcc4 which enhance crystallization. The effect of these domains on the stability and conformation of the myosin rod was examined through biophysical studies and overlapping structures. In addition, a computational approach was developed to combine the sections into a contiguous model. The structures were aligned, trimmed to form a contiguous model, and simulated for >700 ns to remove the discontinuities and achieve an equilibrated conformation that represents the native state. This experimental and computational strategy lays the foundation for building a model for the entire myosin rod. This article is protected by copyright. All rights reserved.