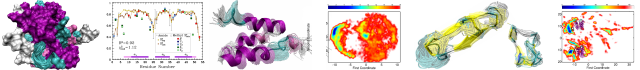

Journal Articles
[Report] Gene essentiality and synthetic lethality in haploid human cells
[Report] Identification and characterization of essential genes in the human genome
[Report] Genome-wide inactivation of porcine endogenous retroviruses (PERVs)
[Report] Cotranslational protein folding on the ribosome monitored in real time
[New Products] New Products
[Podcast] Science Podcast: 27 November Show
[Working Life] The best decision I ever made
CD and NMR investigation of collagen peptides mimicking a pathological Gly–Ser mutation and a natural interruption in a similar highly charged sequence context
Even a single Gly substitution in the triple helix domain of collagen leads to pathological conditions while natural interruptions are suggested to play important functional roles. Two peptides—one mimicking a pathological Gly–Ser substitution (ERSEQ) and the other one modeling a similar natural interruption sequence (DRSER)—are designed to facilitate the comparison for elucidating the molecular basis of their different biological roles. CD and NMR investigation of peptide ERSEQ indicates a reduction of the thermal stability and disruption of hydrogen bonding at the Ser mutation site, providing a structural basis of the OI disease resulting from the Gly–Ser mutation in the highly charged RGE environment. Both CD and NMR real-time folding results indicate that peptide ERSEQ displays a comparatively slower folding rate than peptide DRSER, suggesting that the Gly–Ser mutation may lead to a larger interference in folding than the natural interruption in a similar RSE context. Our studies suggest that unlike the rigid GPO environment, the abundant R(K)GE(D) motif may provide a more flexible sequence environment that better accommodates mutations as well as interruptions, while the electrostatic interactions contribute to its stability. These results shed insight into the molecular features of the highly charged motif and may aid the design of collagen biomimetic peptides containing important biological sites.
Redox-dependent disulfide bond formation in SAP30L corepressor protein: Implications for structure and function
Sin3A-associated protein 30-like (SAP30L) is one of the key proteins in a multi-subunit protein complex involved in transcriptional regulation via histone deacetylation. SAP30L, together with a highly homologous SAP30 as well as other SAP proteins (i.e., SAP25, SAP45, SAP130 and SAP180), is an essential component of the Sin3A corepressor complex, although its actual role has remained elusive. SAP30L is thought to function as an important stabilizing and bridging molecule in the complex and to mediate its interactions with other corepressors. SAP30L has been previously shown to contain an N-terminal Cys3His type zinc finger (ZnF) motif, which is responsible for the key protein-protein, protein-DNA and protein-lipid interactions. By using high-resolution mass spectrometry, we studied a redox-dependent disulfide bond formation in SAP30L ZnF as a regulatory mechanism for its structure and function. We showed that upon oxidative stress SAP30L undergoes the formation of two specific disulfide bonds, a vicinal Cys29-Cys30 and Cys38-Cys74, with a concomitant release of the coordinated zinc ion. The oxidized protein was shown to remain folded in solution and to bind signaling phospholipids. We also determined a solution NMR structure for SAP30L ZnF that showed an overall fold similar to that of SAP30, determined earlier. The NMR titration experiments with lipids and DNA showed that the binding is mediated by the C-terminal tail as well as both α-helices of SAP30L ZnF. The implications of these results for the structure and function of SAP30L are discussed. This article is protected by copyright. All rights reserved.
Characterizing a partially ordered miniprotein through folding molecular dynamics simulations: Comparison with the experimental data
The villin headpiece helical subdomain (HP36) is one of the best known model systems for computational studies of fast-folding all-α miniproteins. HP21 is a peptide fragment –derived from HP36– comprising only the first and second helices of the full domain. Experimental studies showed that although HP21 is mostly unfolded in solution, it does maintain some persistent native-like structure as indicated by the analysis of NMR-derived chemical shifts. Here we compare the experimental data for HP21 with the results obtained from a 15 μs long folding molecular dynamics simulation performed in explicit water and with full electrostatics. We find that the simulation is in good agreement with the experiment and faithfully reproduces the major experimental findings, namely that (a) HP21 is disordered in solution with less that 10% of the trajectory corresponding to transiently stable structures, (b) the most highly populated conformer is a native-like structure with an RMSD from the corresponding portion of the HP36 crystal structure of less than 1Å, (c) the simulation-derived chemical shifts –over the whole length of the trajectory– are in reasonable agreement with the experiment giving reduced χ2 values of 1.6, 1.4 and 0.8 for the Δδ13Cα, Δδ13CO and Δδ13Cβ secondary shifts respectively (becoming 0.8, 0.7, and 0.3 when only the major peptide conformer is considered), and finally, (d) the secondary structure propensity scores are in very good agreement with the experiment and clearly indicate the higher stability of the first helix. We conclude that folding molecular dynamics simulations can be a useful tool for the structural characterization of even marginally stable peptides. This article is protected by copyright. All rights reserved.
Evolution of a Protein Folding Nucleus
The folding nucleus (FN) is a cryptic element within protein primary structure that enables an efficient folding pathway and is the postulated heritable element in the evolution of protein architecture; however, almost nothing is known regarding how the FN structurally changes as complex protein architecture evolves from simpler peptide motifs. We report characterization of the FN of a designed purely-symmetric β-trefoil protein by ϕ-value analysis. We compare the structure and folding properties of key foldable intermediates along the evolutionary trajectory of the β-trefoil. The results show structural acquisition of the FN during gene fusion events, incorporating novel turn structure created by gene fusion. Furthermore, the FN is adjusted by circular permutation in response to destabilizing functional mutation. FN plasticity by way of circular permutation is made possible by the intrinsic C3 cyclic symmetry of the β-trefoil architecture, identifying a possible selective advantage that helps explain the prevalence of cyclic structural symmetry in the proteome. This article is protected by copyright. All rights reserved.
Analysis of free modeling predictions by RBO aleph in CASP11
The CASP experiment is a biannual benchmark for assessing protein structure prediction methods. In CASP11, RBO Aleph ranked as one of the top-performing automated servers in the free modeling category. This category consists of targets for which structural templates are not easily retrievable. We analyze the performance of RBO Aleph and show that its success in CASP was a result of its ab initio structure prediction protocol. A detailed analysis of this protocol demonstrates that two components unique to our method greatly contributed to prediction quality: residue–residue contact prediction by EPC-map and contact-guided conformational space search by model-based search (MBS). Interestingly, our analysis also points to a possible fundamental problem in evaluating the performance of protein structure prediction methods: Improvements in components of the method do not necessarily lead to improvements of the entire method. This points to the fact that these components interact in ways that are poorly understood. This problem, if indeed true, represents a significant obstacle to community-wide progress. Proteins 2015. © 2015 Wiley Periodicals, Inc.
Molecular dynamics of water and monovalent-ions transportation mechanisms of pentameric sarcolipin
The Sarcolipin (SLN) is a transmembrane protein that can form a self-assembled pentamer. In this work, the homology modeling and all-atom molecular dynamic (MD) simulation was performed to study the model of SLN pentamer in POPC (1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine) membrane. The potential of mean force (PMF) was calculated for transmembrane transportation of Na+, Cl− and water molecule along the pore channel of penta-SLN complex. The root mean square deviation (RMSD) of the SLN pentamer in POPC membrane showed that the stabilized SLN protein complex could exist in the membrane and that the Na+ and Cl− could not permeate through the channel when the pore was under the vacuum state, but the water could permeate through from cytoplasm to lumen. Under the aqueous state, our simulation demonstrated that hydrated state of Na+ and Cl− could pass through the channel. The PMF and radii of the pore showed that the channel had a gate at Leu21 that is a key hydrophobicity residue in the channel. Our simulations help to clarify and to understand better the SLN pentamer channel that had a hydrophobic gate and could switch Na+ and Cl− ion permeability by hydrated and vacuum states. Proteins 2015. © 2015 Wiley Periodicals, Inc.
Toward rational thermostabilization of Aspergillus oryzae cutinase: Insights into catalytic and structural stability
Cutinases are powerful hydrolases that can cleave ester bonds of polyesters such as poly(ethylene terephthalate) (PET), opening up new options for enzymatic routes for polymer recycling and surface modification reactions. Cutinase from Aspergillus oryzae (AoC) is promising owing to the presence of an extended groove near the catalytic triad which is important for the orientation of polymeric chains. However, the catalytic efficiency of AoC on rigid polymers like PET is limited by its low thermostability; as it is essential to work at or over the glass transition temperature (Tg) of PET, that is, 70°C. Consequently, in this study we worked toward the thermostabilization of AoC. Use of Rosetta computational protein design software in conjunction with rational design led to a 6°C improvement in the thermal unfolding temperature (Tm) and a 10-fold increase in the half-life of the enzyme activity at 60°C. Surprisingly, thermostabilization did not improve the rate or temperature optimum of enzyme activity. Three notable findings are presented as steps toward designing more thermophilic cutinase: (a) surface salt bridge optimization produced enthalpic stabilization, (b) mutations to proline reduced the entropy loss upon folding, and (c) the lack of a correlative increase in the temperature optimum of catalytic activity with thermodynamic stability suggests that the active site is locally denatured at a temperature below the Tm of the global structure. Proteins 2015. © 2015 Wiley Periodicals, Inc.
Structure and functional analysis of the siderophore periplasmic binding protein from the fuscachelin gene cluster of Thermobifida fusca
Iron acquisition is a complex, multicomponent process critical for most organisms' survival and virulence. Small iron chelating molecules, siderophores, mediate transport as key components of common pathways for iron assimilation in many microorganisms. The chemistry and biology of the extraordinary tight and specific metal binding siderophores is of general interest in terms of host/guest chemistry and is a potential target toward the development of therapeutic treatments for microbial virulence. The siderophore pathway of the moderate thermophile, Thermobifida fusca, is an excellent model system to study the process in Gram-positive bacteria. Here we describe the structure and characterization of the siderophore periplasmic binding protein, FscJ from the fuscachelin gene cluster of T. fusca. The structure shows a di-domain arrangement connected with a long α-helix hinge. Several X-ray structures detail ligand-free conformational changes at different pH values, illustrating complex interdomain flexibility of the siderophore receptors. We demonstrated that FscJ has a unique recognition mechanism and details the binding interaction with ferric-fuscachelin A through ITC and docking analysis. The presented work provides a structural basis for the complex molecular mechanisms of siderophore recognition and transportation. Proteins 2015. © 2015 Wiley Periodicals, Inc.
Towards a Molecular Understanding of the Link between Imatinib Resistance and Kinase Conformational Dynamics
by Silvia Lovera, Maria Morando, Encarna Pucheta-Martinez, Jorge L. Martinez-Torrecuadrada, Giorgio Saladino, Francesco L. Gervasio
Due to its inhibition of the Abl kinase domain in the BCR-ABL fusion protein, imatinib is strikingly effective in the initial stage of chronic myeloid leukemia with more than 90% of the patients showing complete remission. However, as in the case of most targeted anti-cancer therapies, the emergence of drug resistance is a serious concern. Several drug-resistant mutations affecting the catalytic domain of Abl and other tyrosine kinases are now known. But, despite their importance and the adverse effect that they have on the prognosis of the cancer patients harboring them, the molecular mechanism of these mutations is still debated. Here by using long molecular dynamics simulations and large-scale free energy calculations complemented by in vitro mutagenesis and microcalorimetry experiments, we model the effect of several widespread drug-resistant mutations of Abl. By comparing the conformational free energy landscape of the mutants with those of the wild-type tyrosine kinases we clarify their mode of action. It involves significant and complex changes in the inactive-to-active dynamics and entropy/enthalpy balance of two functional elements: the activation-loop and the conserved DFG motif. What is more the T315I gatekeeper mutant has a significant impact on the binding mechanism itself and on the binding kinetics.Reputation Effects in Public and Private Interactions
by Hisashi Ohtsuki, Yoh Iwasa, Martin A. Nowak
We study the evolution of cooperation in a model of indirect reciprocity where people interact in public and private situations. Public interactions have a high chance to be observed by others and always affect reputation. Private interactions have a lower chance to be observed and only occasionally affect reputation. We explore all second order social norms and study conditions for evolutionary stability of action rules. We observe the competition between “honest” and “hypocritical” strategies. The former cooperate both in public and in private. The later cooperate in public, where many others are watching, but try to get away with defection in private situations. The hypocritical idea is that in private situations it does not pay-off to cooperate, because there is a good chance that nobody will notice it. We find simple and intuitive conditions for the evolution of honest strategies.VDJtools: Unifying Post-analysis of T Cell Receptor Repertoires
by Mikhail Shugay, Dmitriy V. Bagaev, Maria A. Turchaninova, Dmitriy A. Bolotin, Olga V. Britanova, Ekaterina V. Putintseva, Mikhail V. Pogorelyy, Vadim I. Nazarov, Ivan V. Zvyagin, Vitalina I. Kirgizova, Kirill I. Kirgizov, Elena V. Skorobogatova, Dmitriy M. Chudakov
Despite the growing number of immune repertoire sequencing studies, the field still lacks software for analysis and comprehension of this high-dimensional data. Here we report VDJtools, a complementary software suite that solves a wide range of T cell receptor (TCR) repertoires post-analysis tasks, provides a detailed tabular output and publication-ready graphics, and is built on top of a flexible API. Using TCR datasets for a large cohort of unrelated healthy donors, twins, and multiple sclerosis patients we demonstrate that VDJtools greatly facilitates the analysis and leads to sound biological conclusions. VDJtools software and documentation are available at https://github.com/mikessh/vdjtools.